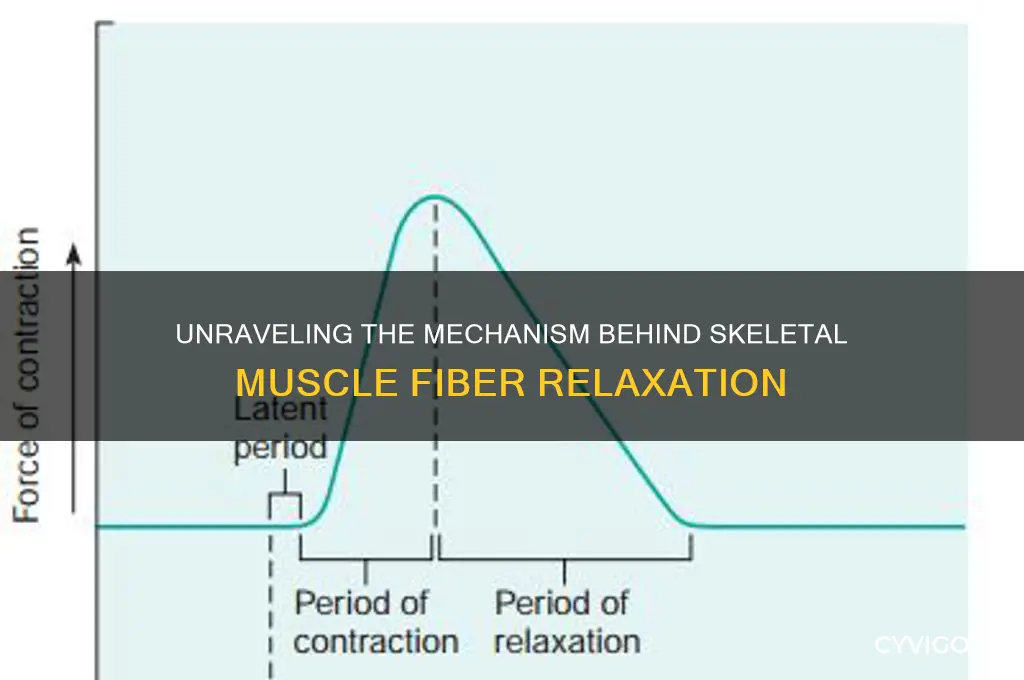
Relaxation in a skeletal muscle fiber is primarily caused by the cessation of calcium ion (Ca²⁺) release from the sarcoplasmic reticulum and the subsequent removal of Ca²⁺ from the cytoplasm. During muscle contraction, Ca²⁺ binds to troponin, exposing myosin-binding sites on actin filaments, allowing cross-bridge formation and muscle shortening. When a nerve signal stops, the muscle fiber’s transverse tubules (T-tubules) cease triggering Ca²⁺ release, and the sarcoplasmic reticulum actively pumps Ca²⁺ back into storage via the calcium ATPase pump. As Ca²⁺ levels drop, troponin reverts to its resting state, blocking myosin-binding sites on actin and preventing further cross-bridge cycling. This process, coupled with the hydrolysis of ATP, detaches myosin heads from actin, allowing the muscle fiber to return to its relaxed state.
| Characteristics | Values |
|---|---|
| Calcium Ion Removal | Calcium ions are actively pumped back into the sarcoplasmic reticulum (SR) by the sarco/endoplasmic reticulum Ca²⁺-ATPase (SERCA) pump, lowering cytoplasmic calcium concentration. |
| Troponin-Tropomyosin Interaction | With reduced calcium, troponin reverts to its resting state, allowing tropomyosin to block myosin-binding sites on actin filaments, preventing cross-bridge formation. |
| Cross-Bridge Detachment | Myosin heads detach from actin filaments due to the absence of calcium-troponin-tropomyosin complex activation, stopping muscle contraction. |
| ATP Hydrolysis | ATP binds to myosin heads, causing them to release actin and return to a high-energy state, ready for the next contraction cycle. |
| Neural Signaling Cessation | Motor neurons stop releasing acetylcholine (ACh), halting action potentials in muscle fibers and ceasing calcium release from the SR. |
| Sarcolemma Repolarization | The muscle fiber membrane repolarizes, closing voltage-gated calcium channels and reducing calcium influx. |
| Role of Nerve Impulses | Absence of nerve impulses leads to reduced calcium release, promoting relaxation. |
| Energy Depletion Prevention | Relaxation conserves ATP, preventing muscle fatigue and allowing for subsequent contractions. |
| Muscle Length Restoration | The muscle returns to its resting length as actin and myosin filaments slide apart. |
| Passive Elastic Elements | Connective tissues and titin (a protein) help restore muscle shape and length during relaxation. |
Explore related products






What You'll Learn
- Calcium Reuptake: Calcium ions return to sarcoplasmic reticulum, ending muscle contraction
- Troponin-Tropomyosin Interaction: Troponin-tropomyosin complex blocks myosin binding sites, stopping contraction
- ATP Hydrolysis: ATP breaks down, releasing energy needed for muscle relaxation
- Sarcomere Structure Reset: Thin and thick filaments return to resting positions
- Neural Signal Cessation: Motor neuron stops releasing acetylcholine, halting muscle stimulation
![]()
Calcium Reuptake: Calcium ions return to sarcoplasmic reticulum, ending muscle contraction
Calcium reuptake is a critical process in the relaxation of skeletal muscle fibers, marking the end of muscle contraction. When a muscle fiber is stimulated to contract, calcium ions (Ca²⁺) are released from the sarcoplasmic reticulum (SR) into the cytoplasm, or sarcoplasm, of the muscle cell. These calcium ions bind to troponin, a protein complex on the actin filaments, causing a conformational change that exposes the myosin-binding sites. This allows myosin heads to attach to actin, initiating the sliding filament mechanism and resulting in muscle contraction. However, for the muscle to relax, this interaction must cease, and calcium reuptake plays a central role in this process.
The termination of muscle contraction begins with the cessation of neural stimulation. When the motor neuron stops releasing acetylcholine at the neuromuscular junction, the muscle fiber’s membrane potential returns to its resting state. This depolarization signal is no longer sustained, leading to the inactivation of voltage-gated calcium channels in the transverse tubules (T-tubules). As a result, the release of calcium ions from the SR into the sarcoplasm is halted. At this point, the existing calcium ions in the sarcoplasm must be actively transported back into the SR to lower their concentration and disrupt the actin-myosin interaction.
Calcium reuptake is facilitated by the sarcoplasmic reticulum calcium ATPase (SERCA) pump, an enzyme embedded in the membrane of the SR. The SERCA pump uses energy from ATP hydrolysis to transport calcium ions against their concentration gradient, from the sarcoplasm back into the SR lumen. This process is highly efficient, rapidly reducing the cytoplasmic calcium concentration to its resting level (approximately 100 nM). As calcium ions are removed from the sarcoplasm, they can no longer bind to troponin, causing the troponin-tropomyosin complex to revert to its inhibitory position. This blocks the myosin-binding sites on the actin filaments, preventing further cross-bridge formation and sliding of the filaments.
The reuptake of calcium ions into the SR is essential for muscle relaxation because it directly reverses the conditions that allowed contraction to occur. Without calcium ions bound to troponin, the actin filaments are effectively shielded from myosin heads, and the muscle fiber returns to its resting state. This mechanism ensures that muscle contraction is a transient event, dependent on the continuous presence of calcium ions in the sarcoplasm. Once calcium reuptake is complete, the muscle fiber is ready to respond to another neural stimulus, maintaining the muscle’s ability to contract and relax efficiently.
In summary, calcium reuptake into the sarcoplasmic reticulum is a fundamental step in the relaxation of skeletal muscle fibers. By actively transporting calcium ions out of the sarcoplasm, the SERCA pump lowers their concentration, disrupting the actin-myosin interaction and terminating contraction. This process is energy-dependent and highly regulated, ensuring that muscle relaxation occurs promptly and completely after the cessation of neural stimulation. Understanding calcium reuptake highlights the intricate interplay between calcium ions, cellular proteins, and energy metabolism in muscle physiology.
Understanding Left Hand Muscle Cramps: Causes and Prevention Tips
You may want to see also
Explore related products






![]()
Troponin-Tropomyosin Interaction: Troponin-tropomyosin complex blocks myosin binding sites, stopping contraction
In skeletal muscle fibers, relaxation is a precisely regulated process that involves the interaction between key proteins, specifically the troponin-tropomyosin complex. This complex plays a critical role in controlling muscle contraction by modulating the interaction between actin and myosin filaments. During muscle relaxation, the troponin-tropomyosin complex acts to block the myosin binding sites on the actin filaments, effectively preventing cross-bridge formation and stopping contraction. This mechanism is central to understanding how muscles transition from a contracted to a relaxed state.
The troponin-tropomyosin complex is composed of three troponin subunits (TnC, TnI, and TnT) and tropomyosin, a long, thin protein that lies in the grooves of the actin filaments. In the absence of calcium ions (Ca²⁺), tropomyosin covers the myosin binding sites on actin, preventing myosin heads from attaching. Troponin I (TnI) binds to actin and holds tropomyosin in this inhibitory position. This structural arrangement ensures that the muscle remains in a relaxed state, as the myosin heads cannot form cross-bridges with actin, and thus, no contraction occurs.
The interaction between troponin and tropomyosin is calcium-dependent, which is a fundamental aspect of muscle relaxation. When a muscle is stimulated to contract, calcium ions are released from the sarcoplasmic reticulum into the cytoplasm. These calcium ions bind to troponin C (TnC), causing a conformational change in the troponin complex. This change leads to the movement of tropomyosin away from the myosin binding sites on actin, exposing them and allowing myosin heads to bind and initiate contraction. Conversely, during relaxation, calcium levels decrease, and the troponin-tropomyosin complex returns to its inhibitory position, blocking the binding sites and halting contraction.
The precise regulation of the troponin-tropomyosin interaction is essential for muscle function. If the complex fails to block the myosin binding sites effectively, the muscle may remain in a partially contracted state, leading to conditions like muscle stiffness or cramps. Conversely, proper functioning ensures that muscles can relax fully, allowing for smooth movement and preventing fatigue. This mechanism highlights the importance of calcium homeostasis and protein interactions in maintaining muscle physiology.
In summary, the troponin-tropomyosin interaction is a key determinant of muscle relaxation in skeletal muscle fibers. By blocking myosin binding sites on actin filaments in the absence of calcium, this complex prevents cross-bridge formation and stops contraction. This process is reversible and tightly regulated by calcium levels, ensuring that muscles can contract and relax efficiently in response to neural signals. Understanding this interaction provides critical insights into the molecular basis of muscle function and relaxation.
TMJ and Chest Pain: Is There a Link?
You may want to see also
Explore related products






![]()
ATP Hydrolysis: ATP breaks down, releasing energy needed for muscle relaxation
ATP hydrolysis plays a pivotal role in the relaxation of skeletal muscle fibers by providing the energy required to reverse the contractile process. When a muscle fiber is stimulated to contract, myosin heads bind to actin filaments, pulling them in a process known as the cross-bridge cycle. This cycle is energetically costly and relies on the energy released from ATP. However, relaxation requires the detachment of myosin heads from actin, a process that also demands energy. ATP hydrolysis is essential here, as it breaks down ATP into ADP (adenosine diphosphate) and inorganic phosphate (Pi), releasing the energy needed to reset the myosin heads to their high-energy state. This energy allows the myosin heads to detach from actin, initiating the relaxation phase.
The process begins when ATP binds to the myosin head, causing it to dissociate from actin. This binding triggers the hydrolysis of ATP, which releases energy and converts ATP to ADP and Pi. The energy from this reaction is used to reposition the myosin head into a "cocked" position, ready for the next contraction cycle. However, during relaxation, this repositioning is crucial for ensuring that the myosin heads remain detached from actin, preventing further sliding of the filaments. Without ATP hydrolysis, the myosin heads would remain bound to actin, leading to sustained contraction or rigidity, a condition known as rigor mortis.
Furthermore, the role of ATP hydrolysis in muscle relaxation is closely tied to calcium ion (Ca²⁺) regulation. During contraction, Ca²⁺ binds to troponin, exposing myosin-binding sites on actin. Relaxation occurs when Ca²⁺ is pumped back into the sarcoplasmic reticulum, lowering its concentration in the cytoplasm. This causes troponin to cover the binding sites on actin, preventing myosin attachment. However, ATP hydrolysis is still necessary to ensure that any remaining myosin heads detach and remain in a non-interactive state. Thus, ATP hydrolysis acts in concert with calcium regulation to ensure complete and efficient muscle relaxation.
Another critical aspect of ATP hydrolysis in muscle relaxation is its role in maintaining the structural integrity of the muscle fiber. The energy released during ATP breakdown is not only used to detach myosin heads but also to restore the sarcomere structure to its resting state. This includes the re-establishment of the overlap between thick and thin filaments, ensuring the muscle is ready for the next contraction. Without this energy, the sarcomeres could remain in a partially contracted state, impairing muscle function and flexibility.
In summary, ATP hydrolysis is indispensable for skeletal muscle relaxation. By breaking down ATP, it releases the energy required to detach myosin heads from actin, reposition them for future contractions, and support calcium-mediated relaxation mechanisms. This process ensures that muscles can transition smoothly from a contracted to a relaxed state, maintaining their responsiveness and preventing stiffness. Understanding ATP hydrolysis highlights its central role in the energetics of muscle function, underscoring its importance in both contraction and relaxation cycles.
Understanding Groin Muscle Spasms: Causes, Triggers, and Prevention Tips
You may want to see also
Explore related products






![]()
Sarcomere Structure Reset: Thin and thick filaments return to resting positions
Relaxation in a skeletal muscle fiber is a highly coordinated process that involves the precise resetting of sarcomere structures. Central to this process is the return of thin and thick filaments to their resting positions, which is essential for muscle relaxation. This reset is driven by the cessation of calcium-mediated activation and the subsequent restoration of the sarcomere’s structural integrity. When a muscle fiber relaxes, the sarcoplasmic reticulum actively pumps calcium ions (Ca²⁺) back into its stores, reducing the cytoplasmic calcium concentration. This removal of calcium from the cytoplasm is critical, as it dissociates calcium from troponin, a protein complex on the thin filaments. Without calcium bound to troponin, the tropomyosin molecules return to their blocking position on the actin filaments, preventing myosin heads from binding to actin. This interruption of cross-bridge formation halts the sliding of thin and thick filaments past each other, effectively stopping muscle contraction.
The return of thin and thick filaments to their resting positions is a passive yet structured process. Thin filaments, composed primarily of actin, tropomyosin, and troponin, revert to their original arrangement along the length of the sarcomere. Tropomyosin covers the myosin-binding sites on actin, ensuring that no further interaction occurs between the filaments. Simultaneously, the thick filaments, composed of myosin molecules, also return to their resting orientation. The myosin heads, no longer bound to actin, adopt a low-energy conformation, ready for the next activation signal. This reset ensures that the sarcomere is prepared for subsequent contractions while maintaining the muscle’s resting length and tone.
The structural reset of the sarcomere is further supported by the elastic properties of titin, a protein that spans the half-sarcomere from the Z-disc to the M-line. Titin acts as a molecular spring, providing passive tension and helping to restore the sarcomere to its resting length during relaxation. As the thin and thick filaments disengage, titin’s elasticity assists in pulling the filaments back to their original positions, maintaining the integrity of the sarcomere structure. This process is crucial for preventing overstretching or damage to the muscle fibers during relaxation.
Additionally, the role of the Z-discs and M-line in sarcomere structure reset cannot be overlooked. The Z-discs, which anchor the thin filaments, act as a stable framework that guides the filaments back to their resting positions. Similarly, the M-line, which anchors the thick filaments, ensures that these filaments remain aligned and organized during relaxation. This coordinated realignment of filaments at the Z-discs and M-line is essential for maintaining the sarcomere’s structural integrity and preparing it for the next contraction cycle.
In summary, the sarcomere structure reset during muscle relaxation is a multifaceted process that relies on the removal of calcium, the passive return of thin and thick filaments to their resting positions, and the supportive roles of proteins like titin, Z-discs, and the M-line. This reset ensures that the muscle fiber is ready for subsequent contractions while maintaining its resting tone and structural stability. Understanding this process provides critical insights into the mechanisms of muscle relaxation and the overall function of skeletal muscle fibers.
Vit D Deficiency: A Cause of Muscle Twitching?
You may want to see also
Explore related products






![]()
Neural Signal Cessation: Motor neuron stops releasing acetylcholine, halting muscle stimulation
Relaxation in a skeletal muscle fiber is a precisely regulated process that begins with the cessation of neural signaling. At the core of this mechanism is the role of the motor neuron and its neurotransmitter, acetylcholine (ACh). When a motor neuron stops releasing acetylcholine into the neuromuscular junction, it initiates a cascade of events that ultimately lead to muscle relaxation. This process, known as Neural Signal Cessation, is fundamental to understanding how skeletal muscles transition from a contracted state to a relaxed state.
The first step in this process occurs at the neuromuscular junction, the specialized synapse between the motor neuron and the muscle fiber. During muscle contraction, the motor neuron releases acetylcholine, which binds to nicotinic acetylcholine receptors (nAChRs) on the muscle fiber’s motor end plate. This binding opens ion channels, allowing sodium ions to flow into the muscle fiber, depolarizing the membrane and triggering an action potential. However, when the motor neuron ceases to release acetylcholine, the stimulation of these receptors stops. Without ACh binding, the nAChRs close, halting the influx of sodium ions and preventing further depolarization of the muscle fiber.
Once the neural signal is discontinued, the muscle fiber’s membrane potential returns to its resting state, a process facilitated by the sodium-potassium pump. This pump actively transports sodium ions out of the cell and potassium ions into the cell, re-establishing the electrochemical gradient. As the membrane potential is restored, the transverse tubules (T-tubules) and sarcoplasmic reticulum (SR) cease their activity in releasing calcium ions (Ca²⁺) into the cytoplasm. Calcium ions are critical for muscle contraction, as they bind to troponin, causing a conformational change that allows myosin heads to interact with actin filaments, generating tension.
With the cessation of acetylcholine release, the lack of new neural signals means no further calcium release from the SR occurs. Additionally, the existing calcium ions in the cytoplasm are actively pumped back into the SR by the calcium ATPase pump. As calcium levels in the cytoplasm decrease, the troponin-tropomyosin complex reverts to its blocking position, preventing myosin heads from binding to actin filaments. This disruption of the actin-myosin interaction is the final step in the relaxation process, as the muscle fiber can no longer sustain contraction.
In summary, Neural Signal Cessation—where the motor neuron stops releasing acetylcholine—is a critical trigger for skeletal muscle relaxation. This cessation halts the depolarization of the muscle fiber, restores the resting membrane potential, and terminates calcium release, ultimately leading to the detachment of myosin heads from actin filaments. This precise and coordinated process ensures that muscles relax efficiently and prepare for the next neural signal, highlighting the elegance of neuromuscular control in the human body.
Neuromuscular Blockade: A Cause of Muscle Pain?
You may want to see also
Frequently asked questions
Relaxation in a skeletal muscle fiber occurs when calcium ions (Ca²⁺) are actively pumped back into the sarcoplasmic reticulum (SR) by the calcium ATPase pump, reducing calcium concentration in the cytoplasm. This prevents calcium from binding to troponin, causing the tropomyosin to block the myosin-binding sites on actin, thus stopping cross-bridge cycling and muscle contraction.
ATP is essential for muscle relaxation because it provides the energy required for the calcium ATPase pump in the sarcoplasmic reticulum to transport calcium ions back into the SR. Additionally, ATP is needed to detach myosin heads from actin during cross-bridge cycling, facilitating the return of the muscle to its relaxed state.
Relaxation is initiated when nerve stimulation ceases, leading to the termination of action potentials in the muscle fiber. Without nerve stimulation, acetylcholine release stops, and the muscle fiber membrane repolarizes. This halts the release of calcium from the sarcoplasmic reticulum, allowing calcium to be actively pumped back into the SR, thereby triggering muscle relaxation.































