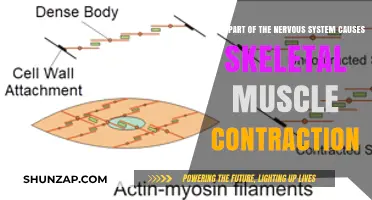
Muscle contractions are primarily driven by a complex interplay of physiological processes involving the nervous system, muscle fibers, and biochemical reactions. At the core of this mechanism is the neuromuscular junction, where motor neurons release acetylcholine, a neurotransmitter that binds to receptors on muscle fibers, initiating an electrical signal. This signal propagates along the muscle fiber’s sarcolemma, triggering the release of calcium ions from the sarcoplasmic reticulum. Calcium ions then bind to troponin, a protein complex on the actin filaments, causing a conformational change that exposes binding sites for myosin heads. The myosin heads attach to actin, pull the filaments past each other, and generate tension, resulting in muscle contraction. This process, known as the sliding filament theory, is powered by ATP hydrolysis, which provides the energy required for myosin’s cyclical interaction with actin. Thus, muscle contractions are a finely orchestrated sequence of electrical, chemical, and mechanical events.
Explore related products






What You'll Learn
- Neural Activation: Motor neurons release acetylcholine, triggering muscle fiber contraction via electrical impulses
- Excitation-Contraction Coupling: Calcium release from sarcoplasmic reticulum binds troponin, enabling actin-myosin interaction
- Sliding Filament Theory: Myosin heads pull actin filaments, shortening sarcomeres and causing muscle contraction
- Energy Metabolism: ATP hydrolysis provides energy for myosin head cycling during muscle contraction
- Muscle Fiber Types: Fast-twitch and slow-twitch fibers contract differently based on myosin isoforms
![]()
Neural Activation: Motor neurons release acetylcholine, triggering muscle fiber contraction via electrical impulses
Muscle contractions are fundamentally driven by neural activation, a process that begins in the central nervous system and culminates in the physical shortening of muscle fibers. At the core of this mechanism are motor neurons, specialized nerve cells that transmit signals from the spinal cord to muscle tissues. When a motor neuron is activated, it propagates an electrical impulse, known as an action potential, down its axon to the neuromuscular junction—the point where the neuron meets the muscle fiber. This electrical signal is the first step in a cascade of events leading to muscle contraction.
At the neuromuscular junction, the arrival of the action potential triggers the release of acetylcholine (ACh), a neurotransmitter stored in vesicles at the terminal end of the motor neuron. Acetylcholine is released into the synaptic cleft, the small gap between the neuron and the muscle fiber. This release is rapid and highly regulated, ensuring precise control over muscle activation. ACh molecules then bind to specific receptors, called nicotinic acetylcholine receptors, located on the surface of the muscle fiber, or sarcolemma. These receptors are ion channels that, when activated, allow sodium ions to flow into the muscle cell, depolarizing the sarcolemma.
The depolarization of the sarcolemma initiates another electrical event within the muscle fiber. This depolarization spreads inward via transverse tubules (T-tubules), which are invaginations of the sarcolemma that penetrate deep into the muscle fiber. The T-tubules are closely associated with the sarcoplasmic reticulum (SR), a network of membranes that stores calcium ions. The depolarization signal triggers the release of calcium ions from the SR into the cytoplasm of the muscle cell, a process mediated by ryanodine receptors on the SR membrane. This sudden increase in intracellular calcium concentration is critical for muscle contraction.
Calcium ions bind to troponin, a protein complex located on the actin filaments of the muscle fiber. This binding causes a conformational change in the troponin-tropomyosin complex, exposing binding sites on the actin filaments. Myosin heads, which are part of the thicker myosin filaments, can then attach to these sites and pull the actin filaments past the myosin filaments, resulting in muscle fiber shortening. This process, known as the sliding filament mechanism, is the basis of muscle contraction. The entire sequence—from neural activation to muscle fiber contraction—is orchestrated by the release of acetylcholine and the subsequent electrical and chemical signaling within the muscle cell.
Finally, the contraction process is terminated by the breakdown of acetylcholine in the synaptic cleft, a task performed by the enzyme acetylcholinesterase. This breakdown ensures that ACh does not continuously stimulate the muscle fiber, allowing for precise control over the duration and intensity of muscle contractions. Simultaneously, calcium ions are actively pumped back into the sarcoplasmic reticulum, lowering the intracellular calcium concentration and allowing the troponin-tropomyosin complex to return to its resting state. This reuptake of calcium prepares the muscle fiber for the next cycle of activation, ensuring that muscle contractions can be repeated as needed for movement and other physiological functions.
Mattress and Muscle Spasms: What's the Link?
You may want to see also
Explore related products






![]()
Excitation-Contraction Coupling: Calcium release from sarcoplasmic reticulum binds troponin, enabling actin-myosin interaction
Muscle contractions are fundamentally driven by a process known as excitation-contraction coupling, which translates electrical signals into mechanical force. This process begins with the arrival of an action potential at the neuromuscular junction, triggering the release of acetylcholine. The action potential then propagates along the muscle fiber's sarcolemma and into the transverse tubules (T-tubules), which are invaginations of the cell membrane. At the junction between the T-tubules and the sarcoplasmic reticulum (SR), known as the triad, voltage-sensing proteins called dihydropyridine receptors (DHPRs) detect the depolarization. This detection initiates a conformational change in the DHPRs, which are physically coupled to ryanodine receptors (RyRs) on the SR membrane.
The activation of RyRs by DHPRs leads to the rapid release of calcium ions (Ca²⁺) from the sarcoplasmic reticulum into the cytoplasm of the muscle cell. This calcium release is a critical step in excitation-contraction coupling. The SR acts as a calcium store, and its release is tightly regulated to ensure precise control of muscle contraction. Once in the cytoplasm, Ca²⁺ ions bind to troponin, a regulatory protein complex located on the thin (actin) filaments of the muscle fiber. Troponin is composed of three subunits: troponin C (TnC), which has a high affinity for calcium, troponin I (TnI), and troponin T (TnT). The binding of Ca²⁺ to TnC induces a conformational change in the troponin-tropomyosin complex.
This conformational change causes tropomyosin, another regulatory protein wrapped around the actin filament, to shift its position. In its resting state, tropomyosin blocks the myosin-binding sites on actin, preventing interaction between the two proteins. However, when calcium binds to troponin, tropomyosin moves away from these binding sites, exposing them. This exposure allows myosin heads (part of the thick filaments) to bind to actin, forming cross-bridges. The formation of these cross-bridges is the first step in the actin-myosin interaction, which is essential for muscle contraction.
The actin-myosin interaction is powered by the hydrolysis of adenosine triphosphate (ATP), the cell's energy currency. Once myosin binds to actin, it undergoes a power stroke, pulling the actin filament toward the center of the sarcomere (the basic contractile unit of muscle). This sliding filament mechanism shortens the sarcomere, leading to muscle contraction. The process is cyclical: myosin releases actin, binds a new ATP molecule, and prepares for the next cycle of binding and contraction. Throughout this cycle, calcium remains bound to troponin, keeping the actin-binding sites accessible.
Termination of muscle contraction occurs when calcium is actively pumped back into the sarcoplasmic reticulum by the sarco(endo)plasmic reticulum Ca²⁺-ATPase (SERCA) pump. As calcium levels in the cytoplasm decrease, it dissociates from troponin, allowing tropomyosin to return to its blocking position on the actin filament. This prevents further myosin binding, and the muscle relaxes. Thus, the release of calcium from the sarcoplasmic reticulum and its binding to troponin are central to enabling the actin-myosin interaction, which is the core mechanism of muscle contraction in excitation-contraction coupling.
Understanding Muscle Mean: Causes, Symptoms, and Effective Treatment Options
You may want to see also
Explore related products






![]()
Sliding Filament Theory: Myosin heads pull actin filaments, shortening sarcomeres and causing muscle contraction
The Sliding Filament Theory is a fundamental concept in understanding muscle contraction, providing a detailed explanation of how muscles generate force and movement. This theory elegantly describes the molecular mechanisms within muscle fibers that lead to contraction. At its core, the process involves the interaction between two key proteins: actin and myosin, which are arranged in a highly organized manner within muscle cells.
In a relaxed muscle, actin and myosin filaments are positioned in a way that allows for potential movement. Actin filaments, also known as thin filaments, are composed of actin proteins and are arranged in parallel arrays, forming a network within the muscle fiber. Myosin filaments, or thick filaments, consist of myosin proteins with their distinctive heads, which can bind to actin. These myosin heads are the driving force behind muscle contraction. When a muscle is stimulated to contract, the myosin heads attach to the actin filaments, initiating a precise sequence of events.
The contraction process begins with the activation of the muscle fiber. Neural signals trigger the release of calcium ions (Ca²⁺) from the sarcoplasmic reticulum, a specialized structure within the muscle cell. This increase in calcium concentration leads to a series of conformational changes in the actin filaments, exposing binding sites for the myosin heads. The myosin heads then undergo a power stroke, pivoting and pulling the actin filaments toward the center of the sarcomere, which is the basic contractile unit of a muscle fiber. This sliding action of the filaments past each other results in the shortening of the sarcomere and, consequently, the entire muscle fiber.
As the myosin heads detach from the actin filaments, they return to their original position, ready for the next cycle of binding and pulling. This cyclic process continues as long as calcium ions remain available, ensuring sustained muscle contraction. The Sliding Filament Theory beautifully illustrates how the precise interaction between actin and myosin, facilitated by calcium-induced changes, leads to the generation of force and muscle movement. This mechanism is essential for various physiological functions, from voluntary movements to maintaining posture and even the beating of the heart.
In summary, the Sliding Filament Theory provides a comprehensive understanding of muscle contraction, highlighting the dynamic interaction between actin and myosin filaments. This theory not only explains the molecular basis of muscle function but also offers insights into the intricate processes that underlie our body's ability to move and perform various physical tasks. By unraveling the mysteries of muscle contraction, scientists and researchers can further explore the complexities of human physiology and potentially develop interventions for muscle-related disorders.
Relapsing Polychondritis: Can It Lead to Muscle Loss?
You may want to see also
Explore related products





![]()
Energy Metabolism: ATP hydrolysis provides energy for myosin head cycling during muscle contraction
Muscle contractions are fundamentally driven by the interaction between actin and myosin filaments, a process that requires a significant amount of energy. This energy is primarily supplied by the hydrolysis of adenosine triphosphate (ATP), the cell's primary energy currency. ATP hydrolysis is a critical step in energy metabolism, providing the necessary power for the myosin head to cycle and generate force during muscle contraction. When ATP binds to the myosin head, it induces a conformational change, allowing the myosin head to detach from actin and move to a new binding site. This cyclical process, known as the cross-bridge cycle, is essential for sustained muscle contraction.
The energy released from ATP hydrolysis is directly utilized to reposition the myosin head, enabling it to bind to a new actin site and pull the filaments past each other. This mechanical action shortens the muscle fiber, resulting in contraction. The efficiency of this process is remarkable, as a single ATP molecule can support multiple myosin head cycles under certain conditions. However, the demand for ATP during intense or prolonged muscle activity is high, necessitating rapid replenishment through various metabolic pathways, including glycolysis, oxidative phosphorylation, and phosphocreatine breakdown.
ATP hydrolysis is not only the energy source for myosin head cycling but also a regulatory mechanism in muscle contraction. The availability of ATP influences the rate and force of contraction. When ATP levels are high, myosin heads can cycle rapidly, leading to stronger and more sustained contractions. Conversely, ATP depletion, as seen in fatigue, slows or halts the cross-bridge cycle, impairing muscle function. This highlights the critical role of energy metabolism in maintaining muscle performance.
The process of ATP hydrolysis is tightly coupled with calcium ion (Ca²⁺) signaling in muscle cells. Ca²⁺ release from the sarcoplasmic reticulum triggers the exposure of myosin binding sites on actin, initiating contraction. ATP hydrolysis then fuels the subsequent mechanical events. This integration of energy metabolism and cellular signaling ensures that muscle contractions are both efficient and responsive to physiological demands. Without ATP, the myosin heads would remain bound to actin, causing muscle stiffness, a condition known as rigor mortis.
In summary, ATP hydrolysis is the cornerstone of energy metabolism in muscle contraction, providing the energy required for myosin head cycling. This process is essential for the dynamic interaction between actin and myosin, enabling muscle fibers to generate force and shorten. The interplay between ATP availability, calcium signaling, and metabolic pathways ensures that muscles can contract effectively under varying conditions. Understanding this mechanism not only sheds light on the physiological basis of muscle function but also underscores the importance of energy metabolism in sustaining physical activity.
Muscle Swelling and Weight Gain: What's the Link?
You may want to see also
![]()
Muscle Fiber Types: Fast-twitch and slow-twitch fibers contract differently based on myosin isoforms
Muscle contractions are fundamentally driven by the interaction between actin and myosin filaments within muscle fibers, a process regulated by neural and biochemical signals. However, not all muscle fibers are created equal. Muscle fibers are broadly categorized into two types: fast-twitch and slow-twitch, each with distinct contractile properties primarily determined by the specific isoforms of myosin heavy chains they express. These myosin isoforms dictate the speed, force, and endurance of muscle contractions, making them central to understanding muscle fiber functionality. Fast-twitch fibers, for instance, contain myosin isoforms that enable rapid contractions, while slow-twitch fibers express isoforms optimized for sustained, efficient contractions.
Fast-twitch muscle fibers, also known as Type II fibers, are characterized by their ability to generate quick, powerful contractions. This is largely due to the presence of myosin isoforms such as MYH2 and MYH4, which have a higher ATPase activity compared to slow-twitch fibers. The higher ATPase activity allows fast-twitch fibers to hydrolyze ATP more rapidly, providing the energy needed for quick, forceful movements. However, this comes at the cost of reduced endurance, as these fibers fatigue more quickly due to their reliance on anaerobic metabolism. Fast-twitch fibers are essential for activities requiring explosive strength, such as sprinting or weightlifting, where speed and power are prioritized over sustained effort.
In contrast, slow-twitch muscle fibers, or Type I fibers, are designed for endurance and sustained contractions. These fibers express myosin isoforms like MYH7, which have lower ATPase activity. This results in slower but more efficient ATP hydrolysis, allowing slow-twitch fibers to maintain contractions over extended periods without fatiguing rapidly. Slow-twitch fibers rely primarily on aerobic metabolism, utilizing oxygen and fatty acids as energy sources, which supports their endurance capabilities. These fibers are crucial for activities like long-distance running or maintaining posture, where sustained, low-intensity contractions are required.
The differences in myosin isoforms between fast-twitch and slow-twitch fibers also influence their response to neural signals. Fast-twitch fibers are innervated by motor neurons with larger diameters, allowing for faster signal transmission and quicker contraction onset. Slow-twitch fibers, on the other hand, are innervated by smaller motor neurons, which conduct signals more slowly but provide greater resistance to fatigue. This neural adaptation complements the biochemical properties of the myosin isoforms, further distinguishing the functional roles of these fiber types.
Understanding the role of myosin isoforms in muscle fiber types has practical implications for training and rehabilitation. Athletes can tailor their training programs to target specific fiber types based on their sport’s demands. For example, sprinters may focus on exercises that activate fast-twitch fibers, while endurance athletes benefit from training that enhances slow-twitch fiber performance. Additionally, this knowledge aids in developing interventions for muscle disorders, as therapies can be designed to address the unique properties of fast-twitch and slow-twitch fibers. In essence, the myosin isoforms in fast-twitch and slow-twitch fibers are the molecular basis for the diverse contractile capabilities of skeletal muscles, shaping their role in movement and performance.
Psoas Muscle and Knee Pain: Unraveling the Hidden Connection
You may want to see also
Frequently asked questions
Muscle contractions are primarily driven by the sliding filament theory, where actin and myosin filaments slide past each other, powered by the hydrolysis of ATP, resulting in muscle fiber shortening.
The nervous system initiates muscle contractions by sending electrical signals (action potentials) through motor neurons, which release acetylcholine at the neuromuscular junction, triggering muscle fiber activation.
Calcium ions (Ca²⁺) bind to troponin in muscle fibers, causing a conformational change that exposes myosin-binding sites on actin, allowing cross-bridge formation and contraction to occur.
ATP provides the energy required for myosin heads to detach from actin filaments and reattach in a new position, enabling the sliding filament process and sustained muscle contraction.
Skeletal muscles contract voluntarily via neural stimulation, smooth muscles contract involuntarily through hormones and neurotransmitters, and cardiac muscles contract rhythmically via intercalated discs and intrinsic pacemaker cells.