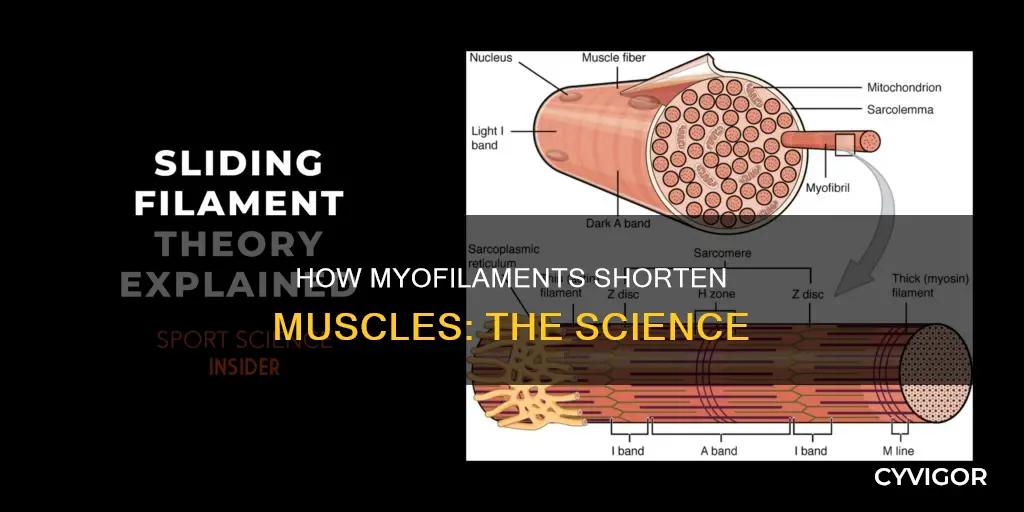
Myofilaments are the three protein filaments of myofibrils in muscle cells. They are responsible for muscle contractions, which occur when the heads of myosin filaments (thick filaments) bind to actin (thin filaments) and pull them past one another, resulting in sarcomere shortening. The thick and thin filaments do not change length during this process, but rather slide past each other, bringing the Z-discs closer together and causing the muscle to contract. This sliding filament theory was first proposed in 1954 and has been a key concept in understanding muscle contraction ever since.
| Characteristics | Values |
|---|---|
| Definition | Myofilaments are the three protein filaments of myofibrils in muscle cells. |
| Types | Thick, thin, and elastic filaments. |
| Composition | Myosin, actin, and titin. |
| Function | Myofilaments work together to cause muscle contraction. |
| Contraction Mechanism | Thick and thin filaments slide past each other, causing the sarcomere to shorten without changing the length of individual filaments. |
| Sarcomere Shortening | The distance between Z-discs reduces, leading to the shortening of the sarcomere and, ultimately, the whole muscle. |
| Contractile Proteins | Myosin and actin. |
| Elastic Protein | Titin. |
| Thick Filament Composition | Myosin, a motor protein. |
| Thin Filament Composition | Actin, tropomyosin, and troponin. |
| Thick Filament Diameter | Approximately 15 nm. |
| Myosin Molecule Shape | Resembles a golf club, with a double globular head and a tail formed of two intertwined chains. |
| Myosin Head Orientation | Half of the myosin heads angle to the left, and half angle to the right, creating a middle area called the M-region or bare zone. |
| Sarcomere Definition | The smallest functional unit of a skeletal muscle fiber, composed of contractile, regulatory, and structural proteins. |
| Sarcomere Length | Increasing length leads to increased force development due to greater thick-thin filament overlap and increased myofilament Ca sensitivity. |
| Inotropic State | Changes in contractile force independent of sarcomere length. Altered by myofilament characteristics, such as kinetics of crossbridge cycling. |
Explore related products






What You'll Learn
![]()
Thick and thin myofilaments
Myofilaments are the three protein filaments of myofibrils in muscle cells. The main proteins involved are myosin, actin, and titin. Myosin and actin are contractile proteins, while titin is an elastic protein. Myofilaments work together during muscle contraction.
Thin myofilaments, on the other hand, are composed primarily of the protein actin, specifically filamentous F-actin. Each F-actin strand is made up of subunits called globular G-actin, which can bind to the head of a myosin molecule. Thin filaments are 7 nm in diameter and contain molecules of tropomyosin, a protein that blocks the active sites of the thin filaments when the muscle is relaxed.
The thick and thin myofilaments are arranged in a specific way within the sarcomere, the repeating unit along the myofibril. The thick myofilaments are anchored at the center of the sarcomere, forming the A band, while the thin myofilaments emerge from the Z disks to form the I bands. The thick and thin filaments interact to generate muscle movement.
The Platysma Muscle: Location and Functionality Explained
You may want to see also
Explore related products






![]()
Myofilament protein changes
Myofilaments are the three protein filaments of myofibrils in muscle cells. The main proteins involved are myosin, actin, and titin. Myosin and actin are contractile proteins, while titin is an elastic protein. The myofilaments work together during muscle contraction, with the thick myosin filament attaching to the thin actin filament and pulling them past one another, resulting in sarcomere shortening and, consequently, muscle contraction.
Research has shown that myofilament protein alterations occur in response to exercise, aging, and disease. While the exact mechanism of myofilament alteration in response to exercise is still being studied, some studies have reported significant differences in mRNA for genes specific to the production of actin. This provides evidence of the mechanisms for both immediate and delayed myofilament responses to exercise at the molecular level. For example, resistance training in humans has been shown to increase specific muscle tension despite a decrease in the presence of MyHC, a myosin heavy-chain. This suggests that there may be no clear relationship between fiber-type composition and in vivo muscle tension.
Additionally, studies on aging have found no alterations in myofilament protein content or isoform expression with age. However, previous studies have suggested a loss of myosin protein content in single muscle fibers from older adults, contributing to the loss of single-fiber tension commonly associated with age.
Furthermore, the presence of disease can also alter myofilament protein content and function. For instance, chronic heart failure (HF) has been shown to cause alterations in myofilament protein content and function, contributing to muscle contractile dysfunction and disability. The type of disease, its etiology, and temporal context all play a role in modifying normal age-related changes in myofilament proteins.
Understanding Muscle Attachment: Hamstring Edition
You may want to see also
Explore related products





![]()
Myosin and actin interaction
Myosin and actin are the contractile proteins that interact to create muscle contractions. Myosin is a motor protein, shaped like a golf club, with a double globular head and a tail formed of two intertwined chains. The actin filaments, on the other hand, are thin and slide past the myosin filaments toward the middle of the sarcomere.
The interaction between these two proteins is often referred to as actomyosin. During muscle contraction, the heads of the myosin filaments attach to the actin filaments, pulling them past one another. This movement results in the shortening of the sarcomere and, therefore, muscle contraction. The actin-myosin interaction produces two types of movements: force generation between actin filaments leading to contractions, and transport of subcellular organelles and macromolecular complexes by myosin motors along actin filaments.
The actin-myosin interaction is regulated by the presence of Ca2+ ions and tropomyosin. When Ca2+ ions are absent, tropomyosin blocks the interaction between myosin and actin. However, when Ca2+ ions are present, tropomyosin moves back into its groove, allowing myosin to move and interact with actin. This regulation is essential for muscle contraction and cellular movement.
The structure of the actin-myosin complex is also important for its function. The actin filaments are linked through a Z-line, with their barbed ends embedded and capped by a protein called 'CapZ'. The pointed ends of the actin filaments are located near the middle of the sarcomere and capped by tropomodulin. The regular spacing between actin filaments is maintained by α-actinin, and the overall structure is stabilised by the giant protein titin.
The Mechanics of Jaw Movement: Muscle Power
You may want to see also
Explore related products

![]()
Sliding filament theory
The sliding filament theory explains the mechanism of muscle contraction based on muscle proteins that slide past each other to generate movement. The sliding filament theory was introduced in 1954 by two research teams working independently of each other. One team consisted of Andrew Huxley and Rolf Niedergerke from the University of Cambridge, and the other team consisted of Hugh Huxley and Jean Hanson from the Massachusetts Institute of Technology. The theory was originally conceived by Hugh Huxley in 1953.
According to the sliding filament theory, the myosin (thick filaments) of muscle fibres slide past the actin (thin filaments) during muscle contraction, while the two groups of filaments remain at a relatively constant length. Myosin and actin form overlapping filaments, with myosin filaments mainly constituting the A band (the dark region of a sarcomere) and actin filaments traversing both the A and I (light region) bands.
The globular heads of myosin bind to actin, forming cross-bridges between the thick and thin filaments. The motor activity of myosin moves its head groups along the actin filament, sliding the actin filaments from both sides of the sarcomere toward the M line, thereby shortening the sarcomere and resulting in muscle contraction. This movement of the actin and myosin filaments past one another brings the Z discs closer together, resulting in the shortening of the sarcomere and, thus, muscle contraction.
The sliding filament theory introduced a new concept called cross-bridge theory, which explains the molecular mechanism of sliding filaments. The cross-bridge theory states that actin and myosin form a protein complex (called actomyosin) by the attachment of the myosin head to the actin filament, forming a cross-bridge between the two filaments.
Muscle Aging: Unraveling the Biology of Strength Loss
You may want to see also
![]()
Myofilament and muscle contraction
Myofilaments are the three protein filaments of myofibrils in muscle cells. The main proteins involved are myosin, actin, and titin. Myosin and actin are contractile proteins, while titin is an elastic protein. Myofilaments work together to cause muscle contraction.
There are three types of myofilaments: thick, thin, and elastic filaments. Thick filaments are primarily composed of myosin, a motor protein. Each thick filament is made up of several hundred molecules of myosin and is approximately 15 nm in diameter. Myosin molecules are shaped like a golf club, with a tail formed of two intertwined chains and a double globular head projecting from it at an angle. Thick filaments are formed by the association of several hundred myosin molecules in a staggered array, with their tails towards the M-line and heads extending toward the Z-discs.
Thin filaments, on the other hand, are primarily composed of actin. Actin filaments are attached to Z-discs at their plus ends and extend towards the center of the sarcomere. The primary component of thin filaments is the actin protein, with two other components being tropomyosin and troponin. Tropomyosin blocks the binding sites and prevents actin-myosin interactions when the muscles are at rest.
Elastic filaments, composed mainly of titin, the largest known protein, help align the thick filaments and add an elastic element to the sarcomere.
During muscle contraction, the myofilaments themselves do not change length. Instead, they slide across each other, causing the distance between the Z-discs to shorten and resulting in the shortening of the sarcomere. This sliding movement is known as the sliding filament theory of muscle contraction. The thick myosin filaments attach to the oppositely oriented thin actin filaments and pull them past one another, resulting in sarcomere shortening and ultimately muscle contraction. The binding of myosin to actin forms cross-bridges that generate filament movement. This movement requires energy, which is provided by ATP.
The inotropic state or contractility refers to changes in contractile force independent of changes in sarcomere length. Altered myofilament characteristics, such as increased sarcomere length and myofilament Ca sensitivity, can affect the inotropic state and the rate of cardiac muscle shortening.
Organs and Muscles: What's the Connection?
You may want to see also
Frequently asked questions
Myofilaments are the three protein filaments of myofibrils in muscle cells. The main proteins involved are myosin, actin, and titin.
During muscle contraction, the myofilaments themselves do not change length, but slide across each other so the distance between the Z-discs shortens, resulting in the shortening of the sarcomere. The thick myofilaments bind with actin, forming cross-bridges between the thick and thin filaments. This sliding of the actin filaments into the A band and H zone results in sarcomere shortening.
A sarcomere is the smallest functional unit of a skeletal muscle fiber and is a highly organized arrangement of contractile, regulatory, and structural proteins. It is the shortening of these individual sarcomeres that leads to the contraction of individual skeletal muscle fibers and ultimately the whole muscle.
There are three different types of myofilaments: thick, thin, and elastic filaments. Thick filaments are composed of myosin protein complexes, while thin filaments are composed of actin.



















