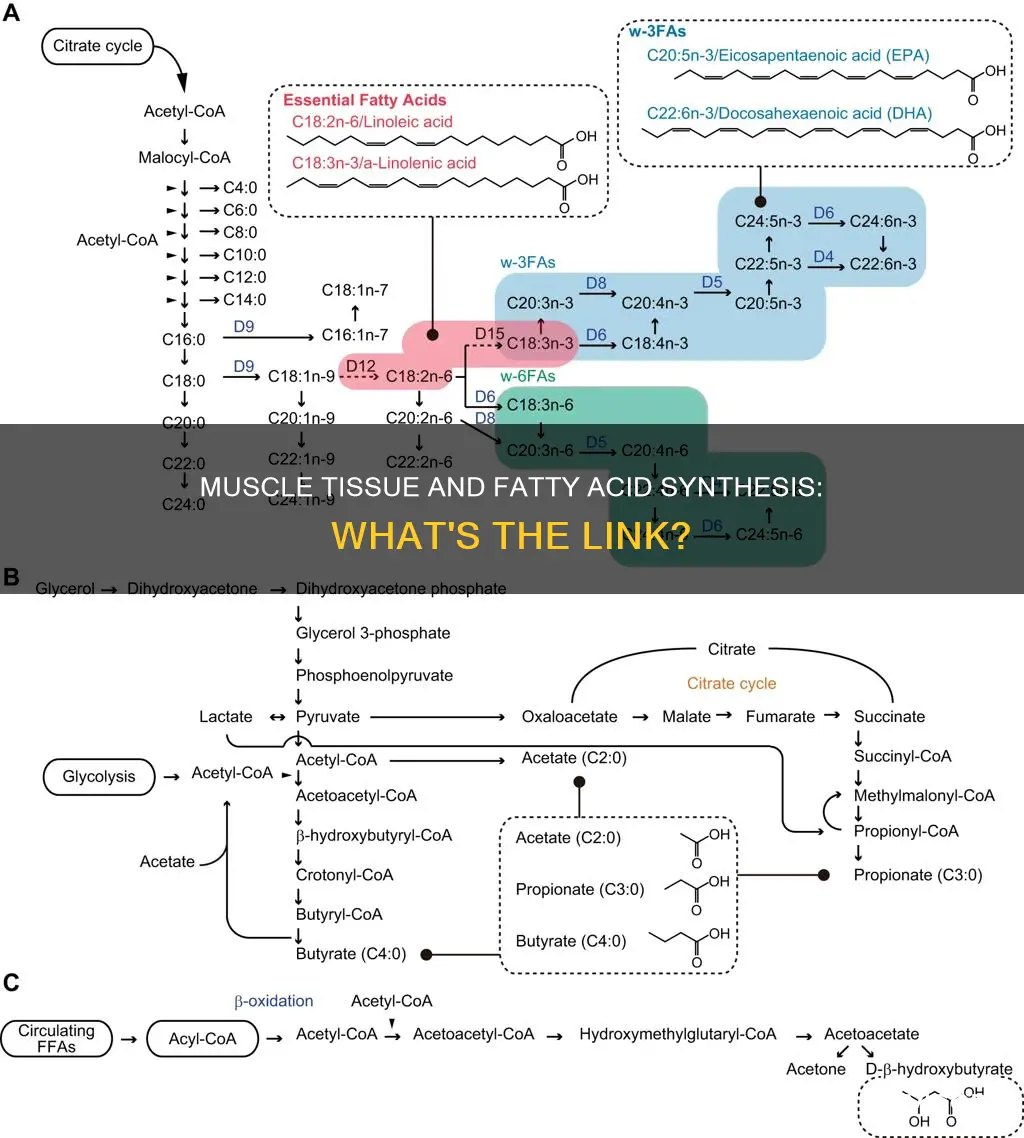
Fatty acids are an important energy source for skeletal muscles. In biochemistry, fatty acid synthesis involves the creation of fatty acids from acetyl-CoA and NADPH through the action of enzymes. The process of fatty acid synthesis can be distinguished into two types: cytosolic fatty acid synthesis (FASI) and mitochondrial fatty acid synthesis (mtFASII). During exercise, triacylglycerols, an energy reservoir in adipose tissue, are hydrolyzed to free fatty acids (FFAs) which are then released into the bloodstream to provide fuel for working muscles.
| Characteristics | Values |
|---|---|
| Fatty acid synthesis | Creation of fatty acids from acetyl-CoA and NADPH through the action of enzymes |
| Fatty acids | An important energy source for skeletal muscle |
| Mitochondrial fatty acid synthesis | Best known for producing lipoic acid |
| Fatty acid oxidation | Required for de novo synthesis of nucleotides in endothelial cells |
| Fatty acid oxidation | The main source of energy for many tissues, including heart and skeletal muscle |
| Fatty acid oxidation | A potential target for anti-angiogenic therapies in cancer |
Explore related products






What You'll Learn
- Fatty acids are an important energy source for skeletal muscle
- Exercise stimulates the release of fatty acids into the bloodstream
- Fatty acid synthesis occurs via six recurring reactions
- Fatty acid oxidation is required for de novo synthesis of nucleotides
- Fatty acid synthesis is the creation of fatty acids from acetyl-CoA and NADPH
![]()
Fatty acids are an important energy source for skeletal muscle
Skeletal muscle cells can utilise fatty acids from various sources. These include non-esterified fatty acids (NEFAs) from circulation and NEFAs originating from the lipolysis of intramuscular triacylglycerol (IMTAG). NEFAs can be taken up by skeletal muscle and directed towards oxidation, esterified into complex lipids, or remain free in the muscle. The fate of NEFAs is influenced by muscle contraction, with contracting muscles exhibiting higher NEFA uptake and directing more NEFAs towards oxidation rather than storage.
The TAG pool in skeletal muscle is in a constant state of flux, with simultaneous synthesis and degradation. During low-intensity exercise, the turnover of the TAG pool increases, indicating that lipolysis is matched by synthesis from the increased uptake of exogenous fatty acids. In contrast, high-intensity exercise leads to a reduction in TAG content due to the repartitioning of incoming fatty acids towards oxidation instead of esterification.
Additionally, skeletal muscle can utilise fatty acids from dietary fat, which reaches the liver, adipose tissue, and skeletal muscle in the form of chylomicron-TAG (CM-TAG). CM-TAG undergoes lipolysis by lipoprotein lipase (LPL), and the resulting NEFAs are taken up by skeletal muscle for oxidation or esterification.
Overall, fatty acids play a crucial role in energy production for skeletal muscle, especially during rest and low-intensity exercise. The balance between fatty acid oxidation and storage is dynamic and influenced by factors such as exercise intensity and muscle contraction.
Color Guard Muscle Memory: Building Physical Strength
You may want to see also
Explore related products






![]()
Exercise stimulates the release of fatty acids into the bloodstream
The TAG pool is in a constant state of flux, with a turnover of 29 hours in resting human skeletal muscle. During low-intensity exercise, TAG turnover increases without changes in TAG content, indicating that lipolysis is matched by synthesis from the increased uptake of exogenous fatty acids. However, during higher-intensity exercise, TAG content is reduced due to the repartitioning of incoming fatty acids away from esterification and towards oxidation.
Hormones such as epinephrine and glucagon stimulate FA release from TG stored in adipocyte lipid droplets, with insulin countering their actions. The stimulating effect of increased epinephrine and decreased insulin levels during exercise leads to an increase in HSL activity and fatty acyl-CoA content in skeletal muscle. This results in the hydrolysis of diacylglycerol (DAG) to a free fatty acid (FFA) and MAG, which is further hydrolyzed by a monoacylglycerol lipase (MGL). The FFAs are then transported to the plasma membrane, leave the adipocyte, and bind to serum albumin in the blood.
Exercise also contributes to an increase in the number of mitochondria in white adipose tissue (WAT) and stimulates the expression of brown adipocyte-specific genes, leading to the "beiging" of WAT and improved glucose intolerance. KLF15, a transcription factor that regulates macronutrient metabolism, is upregulated in skeletal muscle after exercise, playing a crucial role in coordinating fuel utilization. PPARδ, a nuclear receptor, also regulates fatty acid metabolism in muscle, promoting the transformation of muscle fibers to an oxidative type and enhancing fatty acid oxidation.
Alcohol's Impact on Muscles: What You Need to Know
You may want to see also
Explore related products






![]()
Fatty acid synthesis occurs via six recurring reactions
Straight-chain fatty acid synthesis occurs via six recurring reactions, until the 16-carbon palmitic acid is produced. These reactions are performed by fatty acid synthases, which contain multiple enzymes that act as one complex. In animals, these reactions occur on fatty acid synthase I (FASI), a large dimeric protein that has all the enzymatic activities required to create a fatty acid.
Fatty acid synthesis involves the step-by-step addition of two-carbon units to a growing chain. The first step is the carboxylation of acetyl-CoA to malonyl-CoA, followed by the addition of a series of malonyl-CoA units by a complex series of reactions via the multienzyme complex, fatty acid synthase. The carboxylation of acetyl-CoA to malonyl-CoA is catalysed by acetyl-CoA carboxylase, which is the only regulatory enzyme of fatty acid synthesis.
The next step is the transfer of malonyl-CoA to fatty acid synthase. At this point, an acetyl group resides at one site of the enzyme, bound to a sulfhydryl group, while a malonyl group resides at a second site of the enzyme, also bound to a sulfhydryl group. The acyl group at the first sulfhydryl group then attacks the malonyl group at its central carbon. This results in the discharge of CO2 and the elongation of the growing fatty acid chain by two carbons.
The six recurring reactions are performed by fatty acid synthase II (FASII), which is present in prokaryotes, plants, fungi, parasites, and mitochondria. FASII is less efficient than FASI but allows for the formation of more molecules, including medium-chain fatty acids via early chain termination. Once formed, the 16:0 carbon fatty acid can undergo a number of modifications, resulting in desaturation and/or elongation.
Muscle Eddie's Untimely Death: What Happened?
You may want to see also
Explore related products






![]()
Fatty acid oxidation is required for de novo synthesis of nucleotides
Fatty acid oxidation is an important process in muscle metabolism. Skeletal muscle extracts circulating non-esterified fatty acids (NEFA) and fatty acids from intramuscular triacylglycerol (mtag) hydrolysis, which can then be directed towards oxidation, esterified into complex lipids, or remain free in the muscle. The metabolic fate of NEFA entering the muscle is influenced by muscle contraction, with exercise resulting in a higher proportion of NEFA being oxidised rather than stored as mtag. This process of fatty acid oxidation provides an important energy source for skeletal muscle.
Fatty acid synthesis, on the other hand, is the creation of fatty acids from acetyl-CoA and NADPH through the action of enzymes. De novo fatty acid synthesis specifically refers to the synthesis of fatty acids from simple molecules, and it is regulated by two key enzymes: acetyl-CoA carboxylase and fatty acid synthase. Acetyl-CoA carboxylase catalyses the conversion of acetyl-CoA to malonyl-CoA, which is then turned into a fatty acid chain by fatty acid synthase. This process occurs in the cytosol and is essentially the reverse of beta-oxidation, with fatty acids synthesised from successive additions of 2C acetyl-CoA followed by reduction.
While fatty acid oxidation and synthesis are distinct processes, they are interconnected. For example, the by-products of fatty acid metabolism may act as important signals in muscle. Additionally, the oxidation of fatty acids can influence the synthesis of other molecules, such as nucleotides.
Nucleotide synthesis, or de novo nucleotide synthesis, refers to the creation of nucleotides from small precursor molecules such as formate and aspartate. This process does not require the use of free bases such as adenine, guanine, cytosine, thymine, or uracil. Instead, the purine ring is built up atom by atom and attached to ribose, while the pyrimidine ring is synthesised as orotate and attached to ribose phosphate before conversion to common pyrimidine nucleotides.
Fatty acid oxidation is necessary for de novo synthesis because it provides the energy and precursor molecules required for nucleotide synthesis. The oxidation of fatty acids generates acetyl-CoA, which is a crucial molecule for both fatty acid synthesis and nucleotide synthesis. Additionally, the by-products of fatty acid oxidation, such as NADPH, are also required for the synthesis of nucleotides and nucleic acids. Therefore, fatty acid oxidation plays a critical role in providing the energy and substrates necessary for de novo synthesis of nucleotides.
The Heart's Muscular Power: Fact or Fiction?
You may want to see also
Explore related products
![NatureWise CLA 1250 - Conjugated Linoleic Acid Supplement for Women and Men, Not Stimulating, Supports Healthy Weight, Fitness Goals - Non-GMO, Gluten Free - 90 Softgels[1-Month Supply]](https://m.media-amazon.com/images/I/71GShy4NFNL._AC_UL320_.jpg)

$11.6 $19.99




![]()
Fatty acid synthesis is the creation of fatty acids from acetyl-CoA and NADPH
Fatty acid synthesis is the process of creating fatty acids from acetyl-CoA and NADPH through the action of enzymes. This process occurs in two forms: cytosolic fatty acid synthesis (FASI) and mitochondrial fatty acid synthesis (mtFASII). The former involves the use of fatty acid synthase (FASN) polypeptides with multiple domains, while the latter uses separate enzymes.
The synthesis of fatty acids is a complex process that involves multiple reactions and enzymes. In general, acetyl-CoA, derived primarily from carbohydrates via the glycolytic pathway, is converted into fatty acids. The glycolytic pathway also provides glycerol, which combines with three fatty acids through ester bonds to form triglycerides or triacylglycerols, the final product of the lipogenic process. When only two fatty acids combine with glycerol, and the third alcohol group is phosphorylated, a phospholipid is formed. These phospholipids are essential in forming the lipid bilayers that make up cell membranes and surround organelles within cells.
The first committed step in fatty acid synthesis involves converting acetyl-CoA to malonyl-CoA through the action of acetyl-CoA carboxylase. This step is crucial in the synthesis of fatty acids. The subsequent steps involve the successive reduction of the residue, with NADPH acting as a cofactor, resulting in the production of fatty acids. The reducing agent NADPH plays a vital role in fatty acid synthesis, while NAD acts as the oxidizing agent in beta-oxidation, the breakdown of fatty acids back into acetyl-CoA.
Fatty acid synthesis is a dynamic process that occurs in both the cytosol and mitochondria of cells. It is essential for cellular respiration and mitochondrial biogenesis, as well as intracellular signal transduction. The synthesis of fatty acids is influenced by various factors, such as hormonal factors, muscle contraction, and exercise. Additionally, fatty acids synthesized by skeletal muscles can be directed towards oxidation, esterified into complex lipids, or remain free within the muscle.
Understanding Rope Muscle Development: Causes and Prevention
You may want to see also
Frequently asked questions
Fatty acids are created from acetyl-CoA and NADPH through the action of enzymes. In biochemistry, this process is known as fatty acid synthesis. The main fuel stored in the bodies of animals is fat.
Skeletal muscle can extract circulating non-esterified fatty acids (NEFA) and fatty acids originating from the hydrolysis of triacylglycerol transported in plasma lipoproteins. The metabolic fate of plasma NEFA entering the muscle is influenced by muscle contraction, with a higher proportion directed towards oxidation at the expense of storage in mTAG.
Exercise stimulates lipolysis, the hydrolysis of triacylglycerols stored in adipose tissue, resulting in the release of free fatty acids (FFAs) into circulation. During exercise, the mTAG content in contracting muscles decreases, while the uptake of plasma NEFA and the proportion directed towards oxidation increase.