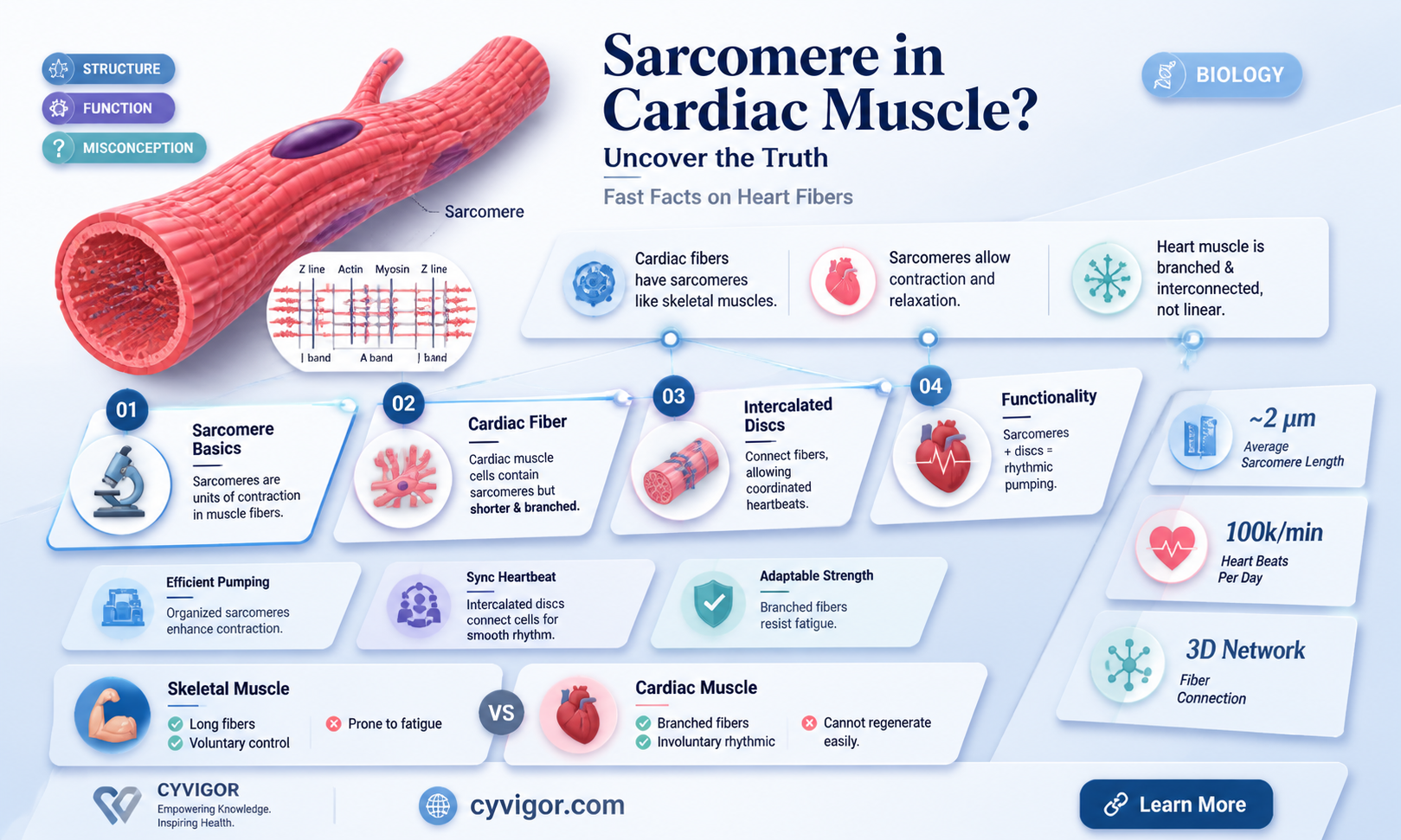
Cardiac muscles, also known as cardiac myocytes, do have sarcomeres. These are the fundamental structural and functional units of striated muscle, generating contractile forces and determining the elastic properties of the muscle. The sarcomere is the region of myofilament structures between two Z-lines and is composed of thick and thin filaments – myosin and actin, respectively. The length of the sarcomere changes as the muscle is stretched, and this change in length is an important mechanism by which the heart regulates its force of contraction.
Explore related products


$12.01 $18.99




What You'll Learn
- Sarcomeres are composed of thick and thin filaments – myosin and actin
- Sarcomere length changes impact the force of cardiac muscle contraction
- The sliding filament theory explains muscle contraction via actin and myosin interaction
- Sarcomeres are responsible for the generation of active and passive forces in cardiomyocytes
- Sarcomeric proteins are the structural building blocks of heart muscle
![]()
Sarcomeres are composed of thick and thin filaments – myosin and actin
The sarcomere is the fundamental unit of striated muscle, including cardiac muscle. It is responsible for the generation of active and passive forces in the heart. Sarcomeres are composed of thick and thin filaments – myosin and actin, respectively. Myosin is a protein with a molecular weight of approximately 470,000 daltons. There are about 300 molecules of myosin per thick filament, and these thick filaments are a key component of sarcomeres. The thick filament mainly consists of myosin molecules, which are part of the myosin superfamily that encodes 18 classes of myosin motors. These myosin motors are ubiquitous in eukaryotes and participate in several cellular motile processes.
Actin, on the other hand, is a globular protein that forms two strands of an alpha helix. It is one of three proteins that make up the thin filaments, the other two being tropomyosin and troponin. Together, these three proteins form what is known as the regulatory protein complex. There are 7 actin molecules per tropomyosin. The actin filaments are attached at their plus ends to the Z-disc, which defines the ends of each sarcomere. The Z-disc also includes the crosslinking protein α-actinin. The thick and thin filaments are connected in the middle of the sarcomere via the M-band.
The interaction between actin and myosin is the basis for the sliding filament theory of muscle contraction. The globular heads of myosin bind to actin, forming cross-bridges between the thick and thin filaments. Myosin contains two heads that serve as the site of the myosin ATPase enzyme, which hydrolyzes ATP required for actin and myosin cross-bridge formation. These cross-bridges are essential for muscle contraction, as they facilitate the sliding of the thick and thin filaments relative to each other, resulting in the shortening of the sarcomere and, consequently, muscle contraction.
The length of the sarcomere, specifically the distance between the Z-lines, is an important factor in regulating the force of contraction. As the sarcomere length increases, so does the force of contraction, or tension development by the muscle fibre. This change in sarcomere length is associated with an increase in the overlap between actin and myosin, providing more sites for cross-bridge formation and, thus, contributing to the increased force of contraction.
Understanding Muscle Stiffness: Causes, Effects, and Remedies
You may want to see also
Explore related products




![]()
Sarcomere length changes impact the force of cardiac muscle contraction
The sarcomere is the fundamental structural and functional unit of striated muscle and is directly responsible for most of its mechanical properties. It is defined as the region of myofilament structures between two Z-lines. In the heart, sarcomeres are responsible for the generation of active and passive forces. The sarcomere length ranges from about 1.6 to 2.2 μm in human hearts.
The sliding filament theory, as described for striated skeletal muscle, proposes that the increased force generation is due to an increase in the overlap between actin and myosin, which makes more sites available for hydrolysis of ATP by the myosin ATPase. However, while this may play a small role in the modulation of active tension development in cardiac muscle, there appear to be other mechanisms that are physiologically more important.
Studies have suggested that the inhibitory region on TnI, including the PKC phosphorylation site (Thr-144), is pivotal for determining the degree of length-dependent augmentation of contractility in cardiac sarcomeres. Replacement of this site with a proline (as in slow skeletal TnI) effectively eliminates myofilament length dependency. Other studies have supported a potential role for titin, the giant elastic protein within each sarcomere, in converting altered passive tension to changes in Ca2+ sensitivity.
Furthermore, changes in preload (the initial length of the muscle before contraction) can affect the force generated by cardiac muscle fibers during isometric contractions (contractions with no change in length) and isotonic contractions (contractions with shortening of the muscle fibers). The changes in active tension caused by changes in preload are related to alterations in the number of actin and myosin cross-bridges formed, which depends on the sarcomere length.
Massage Therapy: Muscle Restructuring and Healing
You may want to see also
![]()
The sliding filament theory explains muscle contraction via actin and myosin interaction
The sliding filament theory, introduced in 1954, explains the mechanism of muscle contraction. It is based on muscle proteins that slide past each other to generate movement. The sliding filament theory is a widely accepted explanation of the mechanism that underlies muscle contraction.
The theory states that the thick filaments of myosin in muscle fibres slide past the thin filaments of actin during muscle contraction. The two groups of filaments remain at a relatively constant length while sliding past each other. This interaction between the actin and myosin filaments generates their movement relative to one another. The molecular basis for this interaction is the binding of myosin to actin filaments, allowing myosin to function as a motor that drives filament sliding.
The sliding filament theory was proposed by two independent research teams. One team consisted of Andrew Huxley and Rolf Niedergerke from the University of Cambridge, and the other team consisted of Hugh Huxley and Jean Hanson from the Massachusetts Institute of Technology. The theory was originally conceived by Hugh Huxley in 1953. The two teams published two consecutive papers on the mechanism in the 22 May 1954 issue of Nature under the common theme "Structural Changes in Muscle During Contraction".
The papers described the position of myosin and actin filaments at various stages of contraction in muscle fibres and proposed how this interaction produced contractile force. Using high-resolution microscopy, the investigators observed changes in the sarcomeres as muscle tissue shortened. They observed that one zone of the repeated sarcomere arrangement, the "A band," remained relatively constant in length during contraction. The A band contains thick filaments of myosin, which suggested that the myosin filaments remained central and constant in length while other regions of the sarcomere shortened. The investigators also noted that the I band, rich in thinner filaments made of actin, changed its length along with the sarcomere.
The sliding filament theory is based on the interaction between actin and myosin. Myosin is a protein with a molecular weight of approximately 470,000 daltons. There are about 300 molecules of myosin per thick filament. Each myosin contains two heads that are the site of the myosin ATPase, an enzyme that hydrolyzes ATP required for actin and myosin cross-bridge formation. These heads interact with a binding site on actin. The thin filaments are composed of three different proteins: actin, tropomyosin, and troponin. Together, these are termed the regulatory protein complex. The actin is a globular protein arranged as a chain of repeating units, forming two strands of an alpha helix. Interdigitated between the actin strands are rod-shaped proteins, termed tropomyosin. There are 7 actin molecules per tropomyosin.
The contraction of myosin's S1 region is called the power stroke. The power stroke requires the hydrolysis of ATP, which breaks a high-energy phosphate bond to release energy. The bending of the myosin S1 region helps explain how myosin moves or "walks" along actin. The slimmer and typically longer "tail" region of myosin (S2) also exhibits flexibility, and it rotates in concert with the S1 contraction. The movements of myosin appear to be a kind of molecular dance. The myosin reaches forward, binds to actin, contracts, releases actin, and then reaches forward again to bind actin in a new cycle. This process is known as myosin-actin cycling.
Exploring the Anatomy of Abdominal Muscles: Unraveling Layers
You may want to see also
![]()
Sarcomeres are responsible for the generation of active and passive forces in cardiomyocytes
The cardiac myocyte is a specialised striated muscle cell, approximately 25μ in diameter and 100μ in length. They form a branching network of cells, connected by intercalated discs. The myocyte is composed of bundles of myofibrils that contain myofilaments. The myofibrils have distinct, repeating microanatomical units, termed sarcomeres, which are the basic contractile units of the myocyte. The sarcomere is the region of myofilament structures between two Z-lines. The sarcomere length, or the distance between Z-lines, ranges from 1.6 to 2.2μ in human hearts.
The thick and thin filaments slide relative to each other, resulting in muscle contraction. Changes in sarcomere length are an important mechanism by which the heart regulates its force of contraction. As a myocyte is stretched, the sarcomeres within the myofibrils are also stretched, and with increased sarcomere length, there is an increase in the force of contraction. This change in length also affects the number of actin and myosin cross-bridges formed, altering the sensitivity of troponin C to calcium, and thus the force of contraction.
Passive tension in the sarcomere is generated by the giant spring protein titin, which is the largest protein in the human proteome. Titin spans the half-sarcomere from the Z-disk to the M-band and stabilises contraction. It also anchors the myosin to the Z-lines. Mutations in sarcomeric proteins, including titin, are responsible for the majority of genetically inherited cardiomyopathies.
Treadmill Training: Friend or Foe to Muscle Gain?
You may want to see also
![]()
Sarcomeric proteins are the structural building blocks of heart muscle
The sarcomere is the fundamental unit of striated muscle, and is directly responsible for most of its mechanical properties. It is composed of thick and thin filaments – myosin and actin, respectively, along with a titin that anchors the myosin to the Z-lines. The cardiac myocyte is a striated muscle cell that forms a branching network of cells that are connected by intercalated discs. The myocyte is made up of bundles of myofibrils that contain myofilaments. The myofibrils contain distinct, repeating microanatomical units, called sarcomeres, which represent the basic contractile units of the myocyte.
The interactions between actin and myosin serve as the basis for the sliding filament theory of muscle contraction. Myosin is a protein with a molecular weight of approximately 470,000 daltons. Each myosin contains two heads that are the site of the myosin ATPase, an enzyme that hydrolyzes ATP required for actin and myosin cross-bridge formation. These heads interact with a binding site on actin. The thin filaments are composed of three different proteins: actin, tropomyosin, and troponin. Together, these are termed the regulatory protein complex.
Changes in sarcomere length are an important mechanism by which the heart regulates its force of contraction. As a myocyte is stretched, the sarcomeres within the myofibrils are also stretched. With increased sarcomere length, there is an increase in the force of contraction. The changes in active tension caused by changes in preload are related to changes in the number of actin and myosin cross-bridges formed, which depends on the sarcomere length.
In the heart, mutations in sarcomeric proteins are responsible for the majority of genetically inherited cardiomyopathies. Sarcomeric dysfunction can lead to cardiac disorders, which represent the main causes of death in Western societies.
Muscle Tension Maintenance: Understanding the Intricate Process
You may want to see also
Frequently asked questions
A sarcomere is the fundamental structural and functional unit of striated muscle. It is composed of thick and thin filaments – myosin and actin, respectively, along with a titin that anchors the myosin to the Z-lines.
Sarcomeres are responsible for the generation of active and passive forces in cardiac muscles. They are directly responsible for most of the mechanical properties of the cardiac muscle.
The sliding filament theory is a theory of muscle contraction. It states that the interactions between actin and myosin, which slide along each other, serve as the basis for muscle contraction.
Mutations in sarcomeric proteins are responsible for the majority of genetically inherited cardiomyopathies. Structural and functional alterations of sarcomeres and their regulators underlie a wide spectrum of different cardiac diseases.
The key components of a sarcomere are actin, myosin, and titin, which make up the thin, thick, and elastic filament systems, respectively.



















